Abstract
Nitrogen stands paramount as a critical nutrient indispensable for wheat growth and productivity. However, nitrogen losses within agroecosystems precipitate diminished soil fertility, decreased crop yield, and environmental perturbations. Ammonia volatilization and denitrification represent primary mechanisms contributing to nitrogen losses in agroecosystems. Consequently, there arises an imperative to explore diverse microbial consortium strategies aimed at enhancing nutrient provision while concurrently mitigating nitrogen losses and aligning with the crop's nutrient demands. In this study, four distinct treatments were implemented: Control (unfertilized, Tc), Azotobacter (Ta), Mycorrhiza (Tm), and Azotobacter + Mycorrhiza (Tam). Notably, the Tam treatment exhibited the highest levels of available soil nutrients such as phosphorus (P) and potassium (K), alongside fostering robust root proliferation, thereby augmenting soil nutrient uptake. Cumulative ammonia flux emissions ranged from 5.84 to 6.25 kg ha-1 in plots receiving microbial consortium treatments, with denitrification losses varying between 3.6 and 3.9 kg ha-1, notably lower compared to Tc-treated plots. Yield assessments revealed a noteworthy yield of 5.34 t ha-1 in the Tam-treated plots, surpassing the yield of Tc-treated plots at 2.73 t ha-1. Consequently, the utilization of microbial consortium practices emerges as an efficacious strategy for curbing nitrogen losses while concurrently enhancing crop yield.
Introduction
Wheat (Triticum aestivum L.) stands as the most extensively cultivated crop worldwide, owing to its remarkable adaptability to diverse environmental conditions and its pivotal role as a staple in the human diet. Of the global wheat production, approximately 65% is allocated for human consumption, with the remainder serving as livestock feed and supporting various facets of the food industry. Notably, wheat serves as the primary source of calories for a significant portion of the global population (Braun et al., 2019). Despite its paramount importance, the rate of yield gain in wheat cultivation has exhibited a continual decline, with yields stagnating in numerous major wheat-producing regions (Fischer and Edmeades, 2020). Climate change coupled with extensive chemical utilization in soil management exacerbates this trend, introducing variability to wheat cultivation conditions and posing challenges to sustainable production intensification. Securing and sustaining future wheat production hinges upon two primary objectives: sustainable intensification of wheat systems to accommodate escalating food demands and comprehending the ramifications of future climate scenarios on wheat yields (Asseng et al., 2011). Nitrogen (N) emerges as a vital nutrient elemental to plant growth and development. However, anthropogenic activities have led to a dramatic surge in the volume of reactive nitrogen (Nr) present in both the biosphere and atmosphere, precipitating a cascade of environmental repercussions including eutrophication, acidification, water pollution, and air pollution (Fowler et al., 2013). Moreover, N losses via various pathways such as runoff, ammonia volatilization, denitrification, and leaching contribute to groundwater pollution, water eutrophication, greenhouse gas emissions, and ozone (O3) degradation in the stratosphere (Shi et al., 2012). Ammonia volatilization, characterized by the release of nitrogen in the form of free ammonia (NH3) gas, stands as a significant pathway for nitrogen losses from agroecosystems. Denitrification, prevalent in agricultural systems, facilitates the conversion of nitrate and nitrite into nitric oxide (NO), nitrous oxide (N2O), and nitrogen gas (N2), with N2 being released under anaerobic conditions (Bouwman et al., 2013). Terrestrial denitrification processes recycle 30%–60% of Nr back into N2 (Ciais et al., 2013), with rice paddy fields or wetlands being identified as major sites of N loss via denitrification (Wang et al., 2017).While nitrogen fertilization stands as a pivotal measure for ensuring high crop yields, its indiscriminate use often leads to problematic agricultural practices. Raiver et al. (2017) observed increases in wheat yield, protein content, and nitrogen use efficiency in response to initial nitrogen deficiencies, potentially attributed to altered root growth dynamics during the seedling stage under low-N conditions. Hence, diversifying nitrogen fertilization strategies beyond reliance on inorganic fertilizers becomes imperative to enhance nitrogen use efficiency. Root elongation and seedling growth in wheat are sluggish during early growth stages, resulting in weak nitrogen absorption capacities owing to immature roots, thereby increasing nitrogen retention or losses and subsequently reducing nitrogen use efficiency (Jiang et al., 2017). Hu et al. (2018) underscored the significance of root morphology and function in nitrogen uptake, with root length, surface area, and branching patterns being primary determinants. Efficient nutrient absorption is facilitated by the upper root system (Tian et al., 2018). Despite achieving self-reliance in agricultural production, the extensive use of chemical fertilizers exacts tolls on soil nutrient statuses and biotic communities. In light of the adverse effects associated with chemical fertilizers, the utilization of microbial consortia as alternative nutrient sources emerges as a promising avenue to meet the escalating demands of crop production sustainably while addressing growing concerns regarding environmental pollution. Microbial consortia, comprising living microorganisms applied to soil, plants, or incorporated with seeds prior to sowing, promote plant growth by enhancing nutrient availability (Dakshayini et al., 2020).
Materials and Methods
The field experiment was conducted at the research farm of the Indian Agricultural Research Institute (IARI), New Delhi, during the winter season of 2018-19. Wheat cultivar "HD 3086" was sown in the third week of November on sandy loam soil with moderate levels of organic carbon (0.45%), nitrogen (347.7 kg/ha), phosphorus (21 kg/ha), and potassium (68.4 kg/ha), accompanied by a soil pH of 8.2 and electrical conductivity of 0.42 dS/m. Meteorological data recorded from the IARI observatory indicated mean minimum and maximum temperatures of 9.18°C and 25.57°C, respectively, throughout the trial period from November to April. The climate of New Delhi is characterized as continental, subtropical, and semi-arid, with an annual average precipitation of 650 mm, primarily attributed to the southwest monsoon.
The experimental design employed a randomized block design with three replications, utilizing wheat variety HD 3086 and four treatments during the winter season: Tc (No Fertilizer), Ta (Azotobacter), Tm (Mycorrhiza), and Tam (Azotobacter + Mycorrhiza). Biofertilizer treatments comprised Mycorrhiza at a rate of 10 kg/ha and Azotobacter seed treatment at 20 kg/ha.
Soil samples were collected from the 0-30 cm soil layer at three distinct locations within each treatment during tillering, flowering, grain-filling, and physiological maturity stages of the crop. Standard procedures were employed to analyze various soil properties, including determination of total nitrogen content using the Kjeldahl method (Kjeldahl, 1883), ammonical nitrogen using continuous flow analyzer, nitrate content using continuous flow analyzer, available soil phosphorus using Olsen’s method (Olsen, 1954), available soil potassium using the Ammonium acetate method, and soil organic carbon using the Walkley and Black method (Walkley and Black, 1934). Plant nitrogen content was estimated via the Kjeldahl method. Ammonia volatilization was assessed using the force air graft method, employing closed chambers (20 cm × 20 cm × 50 cm) constructed from 6 mm acrylic sheets placed in the field. Volatilized ammonia gas was collected in 2% boric acid solution containing a mixed indicator (methyl red and bromocresol green), with air inside the chambers collected into boric acid traps using a vacuum pump operating at a flow rate of 3 L/min. Boric acid traps were replaced every 24 hours. Denitrification potential of soils was determined using the acetylene inhibition method. Harvesting occurred in the fourth week of April 2019, with the recording of yield and yield-contributing characteristics such as number of effective tillers, number of grains per spike, grain yield, and straw yield. Statistical analysis was conducted on the collected data, with means compared at a significance level of 5%.
Results and Discussion
Impact of microbial consortium on soil nutrient status
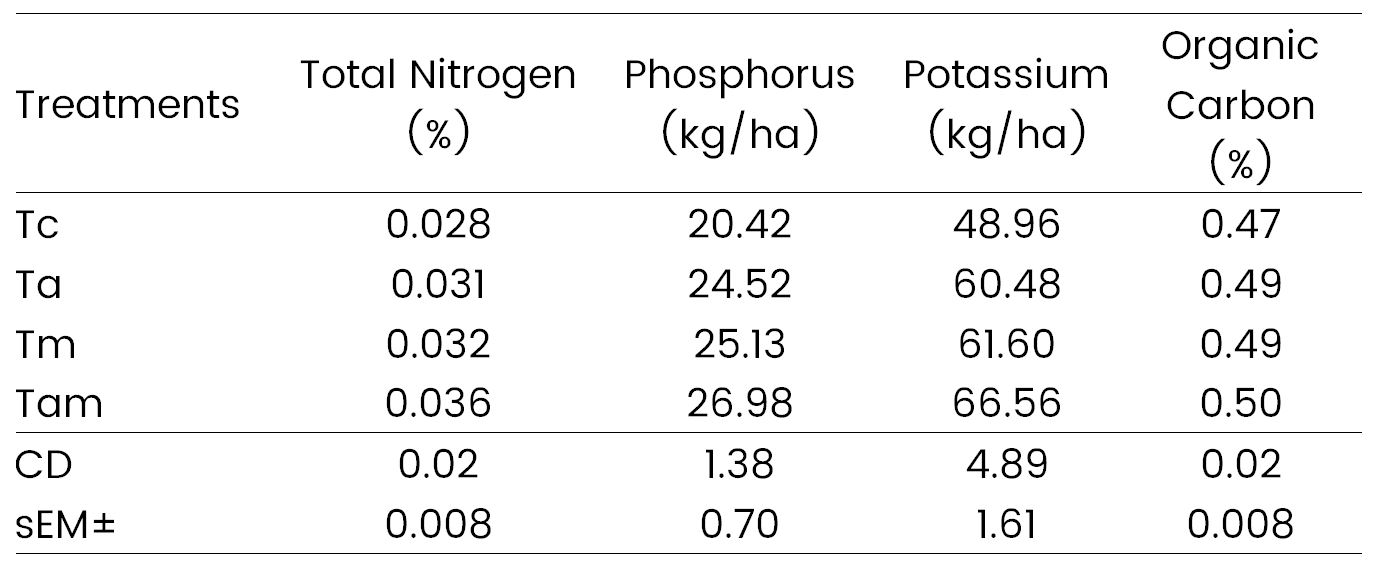
The study revealed significant variations in total nitrogen content among the treatments, ranging from 0.02% to 0.03%. Treatment Tam exhibited the highest total nitrogen content in soil at 0.036%, likely attributable to the application of Azotobacter and Mycorrhiza, resulting in a slight enhancement of total nitrogen content (Table 1). These findings align with those of Karad et al. (2016), who investigated the effect of biofertilizers on nitrogen dynamics in a groundnut-wheat system. Available phosphorus content ranged significantly from 20.4 kg/ha to 26.98 kg/ha, with the highest content observed in treatment Tam. Similarly, available potassium content ranged from 48.9 kg/ha to 66.56 kg/ha, with treatment Tam recording the highest potassium content in soil. Thus, the results indicate that the application of the mycorrhiza biofertilizer in conjunction with Azotobacter enhances soil fertility, thereby contributing to sustainable wheat crop yields. Rama et al. (2015) reported similar findings in soil nutrient studies conducted under various microbial treatments in corn (Zea mays L.). Notably, regular addition of biofertilizers in combination resulted in a significant improvement in soil organic carbon content, ranging from 0.47% to 0.50%, with treatment Tam exhibiting the highest organic carbon content at 0.50%. This increase in organic carbon content may be attributed to enhanced soil microbial diversity associated with wheat crops. Kaur et al. (2018) also observed similar results in their study on soil organic matter dynamics influenced by long-term organic and inorganic fertilizer use in a maize-wheat cropping system.
Table 1. Effect of microbial consortium on soil nutrient status
Impact of microbial consortium on Nitrogen losses
The investigation revealed significant reductions in ammonium volatilization and denitrification losses in plots receiving Azotobacter and Mycorrhiza. The lowest losses were observed in treatment Tc at 3.8 N kg/ha, while treatment Tam recorded the highest loss at 6.2 N kg/ha. Treatments Ta and Tm also exhibited decreased ammonium volatilization losses, with treatment Tam statistically comparable to treatments Tr and Ta. Similarly, for denitrification losses, treatment Tam recorded the highest loss at 3.9 N kg/ha, whereas the lowest losses were observed in treatment Tc at 1.05 N kg/ha. Treatments Ta and Tm demonstrated a two-fold decrease in denitrification losses, with treatments Ta and Tm statistically comparable in denitrification losses (Fig. 1). This reduction in denitrification losses under integrated fertilizer treatment compared to inorganic fertilizer treatment aligns closely with the findings of Aulakh (2020) on the use of biofertilizers for sustainable crop production, improved crop quality, soil health, and reduced environmental pollution.
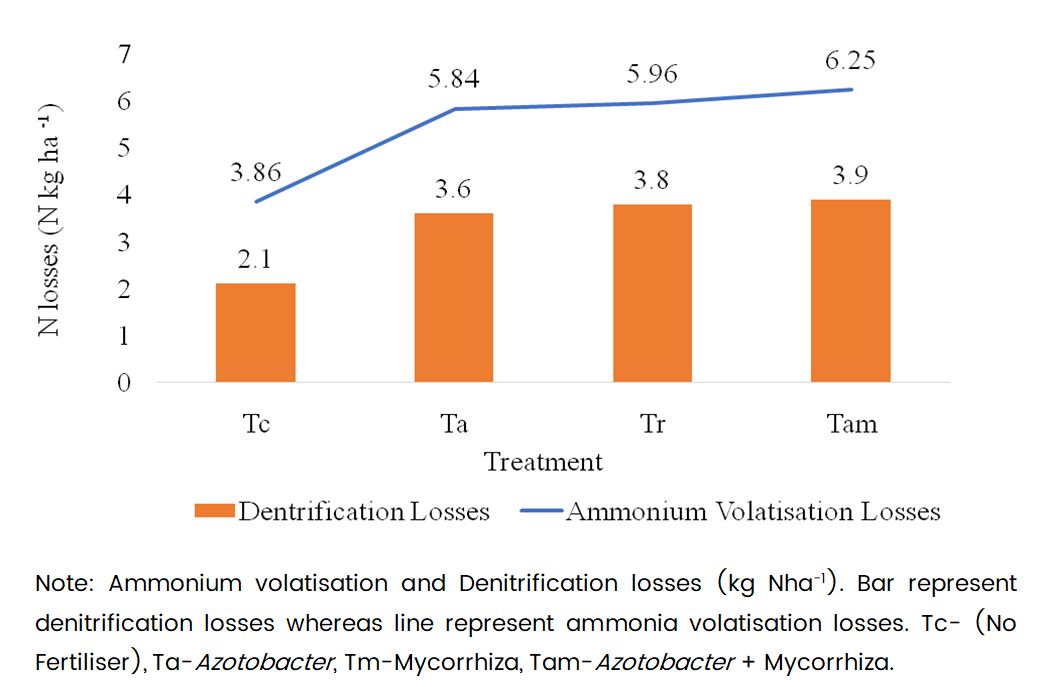
Fig. 1 Effect of microbial consortium on Nitrogen losses
Impact of microbial consortium on Wheat crop yield attributes
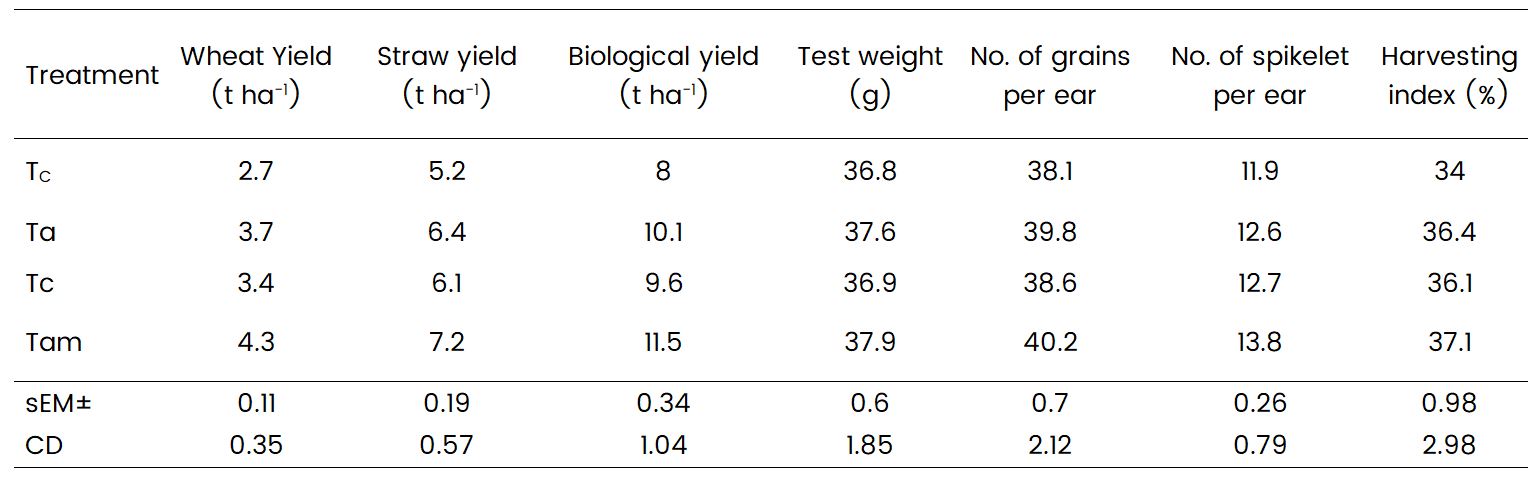
Azotobacter and Mycorrhiza significantly influenced wheat yield attributes, including grain yield, straw yield, test weight, number of grains per ear, and number of spikelets per ear (Table 2). Treatment Tam exhibited the highest grain yield at 4.3 t/ha, significantly higher compared to treatments Tr and Ta, with a percentage increase of 4.7% compared to Tr and Ta. Treatment Tam also demonstrated the highest test weight at 37.9 g, representing a 5.9% increase compared to treatment Tc. Treatment Tam also recorded the highest number of grains per ear and number of spikelets per ear at 40.2 and 13.8, respectively. The harvesting index was highest in treatment Tam at 37.1%, significantly higher compared to treatments Tr and Tc, with treatments Ta and Tr exhibiting statistically comparable values. Significant differences in yield were observed in treatments receiving biofertilizer compared to the unfertilized control, consistent with the findings of
Soleimanzadeh and Gooshchi (2018) on the effects of Azotobacter and nitrogen chemical fertilizer on wheat yield and yield components. The increase in grain and straw yield may be attributed to the sufficient quantities and balanced fractions of plant nutrients delivered to the crop as needed during the growth period, resulting in enhanced yield attributes. The integrated use of fertilizers with biofertilizers promoted dry matter accumulation, increased the number of effective tillers, grains per spike, and test weight, likely due to stimulated vegetative growth facilitated by the sustained and adequate supply of essential nutrients, as reported by Devi et al. (2019) in their study on the effect of microbial consortium on wheat growth and yield.
Table 2: Effect of microbial consortium on yield attributes of wheat crop
Conclusion
The findings of this study demonstrate that the application of biofertilizers enhances the pool of available nutrients in the soil, thereby increasing their accessibility for plant uptake. Moreover, it improves nitrogen use efficiency, rendering the soil less prone to nitrogen losses, whether through volatilization or denitrification. This was evidenced by the reduction in rates of ammonia volatilization and denitrification losses with the utilization of Azotobacter and Mycorrhiza treatments in wheat cultivation. Additionally, the study highlights the positive impact of Azotobacter and Mycorrhiza on wheat yield attributes, indicating the agronomic feasibility, economic viability, and environmental sustainability of employing these microbial treatments in crop production systems. By augmenting soil fertility and mitigating nitrogen losses, the integration of both nutrient sources optimizes fertilizer inputs, enhances crop nutrient use efficiency, and ultimately reduces nutrient losses, thereby contributing to sustainable crop production practices.
References
Asseng S, Foster I A N and Turner N C. 2011. The impact of temperature variability on wheat yields. Global Change Biology 17(2): 997-1012. Doi: http://dx.doi.org/10.1111/j.1365-2486.2010.02262.x
Aulakh M S. 2020. Integrated nutrient management for sustainable crop production, improving crop quality and soil health, and minimizing environmental pollution. In 19th world congress of soil science, soil solutions for a changing world. pp. 1-6.
Bouwman A F, Beusen A H W, Griffioen J, Van Groenigen J W, Hefting M M, Oenema O and Stehfest E. 2013. Global trends and uncertainties in terrestrial denitrification and N2O emissions. Philosophical Transactions of the Royal Society B: Biological Sciences 368(1621). Doi: doi.org/10.1098/rstb.2013.0112
Braun H J, Atlin G and Payne T. 2019. Multi-location testing as a tool to identify plant response to global climate change. Climate change and crop production 1: 115-138.
Ciais P, Sabine C, Bala G, Bopp L, Brovkin V, Canadell J, Chhabra A, DeFries R, Galloway J, Heimann M, Jones C, Quere C L, Myneni R B, Piao S and Thornton P. 2013. Carbon and other biogeochemical cycles. In Climate change 2013: the physical science basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change, Cambridge University Press. pp. 465-570.
Dakshayini G, Anantha Rama A, Sandhya GM, Narayanaswamy B and Muthuraju R. 2020. Screening of efficient phosphate solubilising bacteria from different chickpea growing areas of Karnataka. Journal of Pharmacognosy and Phytochemistry 9(5): 2566-2570. DOI: 10.22271/phyto.2020.v9.i5aj.12728
Devi K N, Singh M S, Singh N G and Athokpam H S. 2019. Effect of integrated nutrient management on growth and yield of wheat (Triticum aestivum L.). Journal of Crop and Weed 7(2): 23-27.
Fischer R A and Edmeades G O. 2012. Breeding and cereal yield progress. Crop Science 50: S-85. Doi: https://doi.org/10.2135/cropsci2009.10.0564
Fowler D, Coyle M, Skiba U, Sutton M A, Cape J N, Reis S, Sheppard L J, Jenkins A, Galloway J N, Vitousek P, Leech A, Bouwman A F, Butterbach-Bahl K, Dentener F, Stevenson D, Amann M and Voss M. 2013. The global nitrogen cycle in the 21st century. Philosophical Transactions of the Royal Society Ser. B. 368(1621): 20130164. Doi: https://doi.org/10.1098/rstb.2013.0164
Hu C, Tian Z, Gu S, Guo H, Fan Y, Abid M, Chen K, Jiang D, Cao W and Dai T. 2018. Winter and spring night-warming improve root extension and soil nitrogen supply to increase nitrogen uptake and utilization of winter wheat (Triticum aestivum L.). European Journal of Agronomy 96: 96-107. Doi: https://doi.org/10.1016/j.eja.2018.03.008
Jiang S, Sun J, Tian Z, Hu H, Michel E J, Gao J and Dai T. 2017. Root extension and nitrate transporter up-regulation induced by nitrogen deficiency improves nitrogen status and plant growth at the seedling stage of winter wheat (Triticum aestivum L.). Environmental and Experimental Botany 141: 28-40.
Karad G U, Babariya N B, Viradiya M B, Parmar J K, Deshmukh S D and Polara K B. 2016. Effect of integrated nutrient management on yield and active pools of soil organic carbon under groundnut-wheat system of typic Haplustept in long term fertilizer experiment. Journal of Pure and Applied Microbiology 10(1): 787-794.
Kaur T, Brar B S and Dhillon N S. 2018. Soil organic matter dynamics as affected by long-term use of organic and inorganic fertilizers under maize–wheat cropping system. Nutrient Cycling in Agroecosystems 81(1): 59-69. Doi: https://doi.org/10.1007/s10705-007-9152-0
Kjeldahl C. 1883. A new method for the determination of nitrogen in organic matter. Z Analytical Chemistry 22: 366.
Olsen S R. 1954. Estimation of available phosphorus in soils by extraction with sodium bicarbonate (No. 939). US Department of Agriculture.
Rama A A, Mahadevaswamy K, Naik N and Kuruber A R. 2015. Influence of Efficient Strain of PSB on Growth and Yield of Maize (Zea mays L.) Under Black Cotton Soil Condition. Journal of Pure and Applied Microbiology 9(2): 1179-1184.
Ravier C, Meynard J M, Cohan J P, Gate P and Jeuffroy M H. 2017. Early nitrogen deficiencies favor high yield, grain protein content and N use efficiency in wheat. European Journal of Agronomy 89: 16-24. Doi: https://doi.org/10.1016/j.eja.2017.06.002
Shi Z, Jing Q, Cai J, Jiang D and Cao W. 2012. The fates of 15N fertilizer in relation to root distributions of winter wheat under different N splits. European Journal of Agronomy 40: 86-93. Doi: https://doi.org/10.1016/j.eja.2012.01.006
Soleimanzadeh H and Gooshchi F. 2018. Effects of Azotobacter and nitrogen chemical fertilizer on yield and yield components of wheat (Triticum aestivum L.). World Applied Sciences Journal 21(8): 1176-1180.
Tian Z W, Liu X X, Gu S L, Yu J H, Zhang L, Zhang W W, Jiang D, Cao W-X and Dai T-B. 2018. Postponed and reduced basal nitrogen application improves nitrogen use efficiency and plant growth of winter wheat. Journal of Integrative Agriculture 17(12): 2648-2661. Doi: https://doi.org/10.1016/S2095-3119(18)62086-6
Walkley A and Black I. A. 1934. An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Science 37(1): 29-38.
Wang S, Shan J, Xia Y, Tang Q, Xia L, Lin J and Yan X. 2017. Different effects of biochar and a nitrification inhibitor application on paddy soil denitrification: a field experiment over two consecutive rice-growing seasons. Science of the Total Environment 593: 347-356. Doi: https://doi.org/10.1016/j.scitotenv.2017.03.159
Copyright
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.

